| Journal of Hematology, ISSN 1927-1212 print, 1927-1220 online, Open Access |
| Article copyright, the authors; Journal compilation copyright, J Hematol and Elmer Press Inc |
| Journal website https://jh.elmerpub.com |
Original Article
Volume 15, Number 2, April 2026, pages 108-128

Engraftment Outcome of CRISPR/Cas9-Edited Hematopoietic Stem Cells for Genetic Diseases: A Systematic Review and Meta-Analysis of Preclinical Evidence
Sudhanshu Yadava, g, Bandana Chakravartib, Baby Anjumc, Shubhanshu Yadavd, Prashant Kumar Singhe, Ashok Kumarf
aStem Cell Research Centre, Department of Hematology, Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow, Uttar Pradesh 226014, India
bStem Cell/Cell Culture Lab Unit, Centre for Advance Research, King George’s Medical University, Lucknow 226003, India
cDepartment of Neurology, Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow 226014, India.
dIQVIA, Marathahalli, Bengaluru 560103, India
eDepartment of Biochemistry, University of Lucknow, Lucknow 226007, India
fDepartment of Biochemistry and Biophysics, Rochester University Medical Centre, Rochester, NY 14625, USA
gCorresponding Author: Sudhanshu Yadav, Stem Cell Research Centre, Department of Hematology, Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow, Uttar Pradesh 226014, India
Manuscript submitted February 2, 2026, accepted March 11, 2026, published online April 6, 2026
Short title: CRISPR/Cas9-Edited HSPC and Engraftment
doi: https://doi.org/10.14740/jh2190
| Abstract | ▴Top |
Background: CRISPR-Cas9 (clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9)-based gene editing represents a promising frontier for treating monogenic hematologic disorders. Several preclinical studies have demonstrated the transplantation efficiency of CRISPR-Cas9-mediated gene editing in hematopoietic stem and progenitor cells (HSPCs) using various animal models. Nonetheless, these studies have employed diverse gene-editing strategies, utilizing HSPCs from different origins and transplanting them into distinct mouse strains. The present study aimed to determine the optimum conditions for efficient engraftment of genetically modified HSPCs across various organs, thereby facilitating the translation of preclinical research into clinical applications.
Methods: We conducted a comprehensive literature search using PubMed Medline, Web of Science, and Google Scholar for relevant articles published from 2014 to 2025 that evaluated the engraftment potential of CRISPR-Cas9 HSPCs in genetic disease models. A total of 39 studies met the inclusion criteria and were included in a meta-analysis using Jamovi software.
Results: The study revealed a significantly reduced engraftment of gene-edited cells in the bone marrow, spleen, and peripheral blood in the pooled analysis. Subgroup analyses revealed that knockout cells exhibited diminished engraftment, whereas knock-in cells demonstrated engraftment levels comparable to those of their non-edited counterparts. No evidence of publication bias or substantial heterogeneity in the study design or outcomes was detected.
Conclusions: Identifying the optimal parameters for gene editing to enhance engraftment efficiency may provide crucial insights for designing future clinical trials and advancing the therapeutic application of CRISPR-Cas9 edited HSPCs.
Keywords: Hematopoietic stem cell; Gene editing; Transplantation; Mice model; Engraftment efficiency; Peripheral blood; Bone marrow; Spleen
| Introduction | ▴Top |
Genetic diseases from mutations in hematopoietic cell lineages, such as sickle cell disease, β-thalassemia, and severe combined immunodeficiency (SCID), pose significant clinical challenges with limited curative options available. Hematopoietic stem cell transplantation (HSCT) offers a potential cure for numerous inherited conditions, including thalassemia, bone marrow (BM) failure syndromes, primary immunodeficiencies, Gaucher disease, Hunter syndrome, or various mucopolysaccharidoses [1]. In HSCT, hematopoietic stem cells are intravenously infused to restore blood cell production. Transplants can be autologous (patient’s own cells), allogeneic (from a donor), or syngeneic (from an identical twin). Cell sources include BM, peripheral blood (PB), umbilical cord blood (UCB), and fetal liver, each with distinct advantages and limitations. Successful transplantation depends on donor availability and conditioning regimens involving chemotherapy, radiation, or monoclonal antibodies [2]. Currently, HSCT is used to treat over 70 diseases, with the number growing rapidly. According to the Worldwide Network for Blood and Marrow Transplantation, approximately 90,000 HSCTs are performed annually worldwide, of which 53% are autologous and 47% are allogeneic [3]. Advances in conditioning strategies, infection control, and graft-versus-host disease (GVHD) management have improved patient outcomes despite persistent donor matching challenges [4].
Gene therapy has emerged as a transformative approach for treating genetic disorders, particularly those involving hematopoietic dysfunctions. Ex vivo gene transfer into hematopoietic stem and progenitor cells (HSPCs) is a pivotal strategy for diseases amenable to BM transplantation [5]. Hematopoietic gene therapy has pioneered the initial clinical proof of concept for treating SCID in human patients [6]. Recent advancements in genome-editing technologies such as transcription activator-like effector nucleases (TALENs), zinc-finger nucleases (ZFNs), homing endonucleases, and especially CRISPR-Cas9 (clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9) facilitate precise, efficient, and versatile modifications of cellular genomes [7]. When HSPCs are edited ex vivo and subsequently transplanted, they can achieve long-term engraftment and confer systemic therapeutic benefits [8]. Currently, several hematopoietic gene therapy products are available, including the first CRISPR/Cas9-based therapy using ex vivo edited HSPCs for treating sickle cell disease [9].
The success of hematopoietic gene therapy has historically relied on myeloablative conditioning, which enhances multilineage HSPC engraftment and enables sustained gene correction [10]. Over the years, mobilized PB HSPCs have emerged as the standard cell source for ex vivo therapies, demonstrating long-term multilineage engraftment comparable to that of BM-derived HSPCs [11]. Gene transfer vectors play a crucial role in ensuring the safety and efficacy of these therapies [12]. Recent advances, including mRNA-based and non-viral in vivo approaches, offer the potential for non-genotoxic conditioning, thereby obviating the need for chemotherapy by facilitating the selection of HSPCs with a high degree of gene modification [13]. Despite these advancements, a significant challenge persists in achieving a safe gene dose that ensures an effective therapeutic level in vivo [14]. Preclinical evaluation of gene-edited HSPCs in animal models is essential to assess engraftment efficiency, long-term persistence, and functional correction of target diseases [15]. Engraftment reflects the ability of edited HSPCs to repopulate the BM and sustain multilineage hematopoiesis [16]. Despite numerous studies demonstrating CRISPR/Cas9-mediated gene editing in HSPCs, variability in design, delivery, efficiency, and outcome metrics makes it challenging to synthesize the data. This systematic review and meta-analysis consolidates current preclinical evidence to quantitatively assess the engraftment potential of CRISPR/Cas9-edited HSPCs in curing monogenic hematologic disorders.
| Materials and Methods | ▴Top |
Study design
We conducted this study in accordance with the protocols established in previously published systematic reviews and meta-analysis, adhering to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [17].
Eligibility criteria
We established specific inclusion and exclusion criteria for the results obtained from the literature search and screened the studies accordingly. The inclusion criteria were as follows: 1) original and full-length articles; 2) preclinical animal models transplanted with CRISPR/Cas9-edited HSPCs; 3) non-edited HSPCs; 4) studies using laboratory mice; and 5) articles published in English.
The exclusion criteria were as follows: 1) review articles; 2) clinical reports and/or trials; 3) studies involving non-CRISPR gene editing techniques; and 5) studies that failed to provide the required information. There were no restrictions regarding species, age, gender, duration of transplantation, and administration of CRISPR/Cas9-edited HSPCs.
Search strategy
Three independent researchers (SY, PKS, and AK) conducted a comprehensive literature search using PubMed Medline, Web of Science, and Google Scholar to identify studies that assessed the effect of CRISPR/Cas9-mediated gene editing of HSPCs on transplantation in mice to correct genetic diseases. The search strategy employed various terms, including (“CRISPR-Cas9”) AND (“gene therapy” OR “gene editing”) AND (“Hematologic disorder” OR “Hemoglobinopathy” OR “Hemoglobinopathies” OR “sickle cell disease” OR “sickle cell anemia” OR “SCD” OR “hematopoietic progenitor stem cell” OR “HSPC” OR “hematopoietic stem cell” OR “HSC” OR “thalassemia” OR “beta thalassemia” OR “alpha thalassemia” OR “thalassemia major” OR “thalassemia minor” OR “animal engraftment” OR “mouse engraftment” OR “HSC engraftment” OR “HPSC engraftment” OR “stem cell transplantation” OR “bone marrow transplantation” OR “peripheral blood stem cell transplantation” OR “hematopoietic stem cell transplantation” OR “HSC transplantation” OR “hematopoietic progenitor stem cell transplantation” OR “HPSC transplantation”). We manually reviewed the references cited in the relevant articles. The results of the literature search are presented in a PRISMA flow chart (Fig. 1), showing the study selection process.
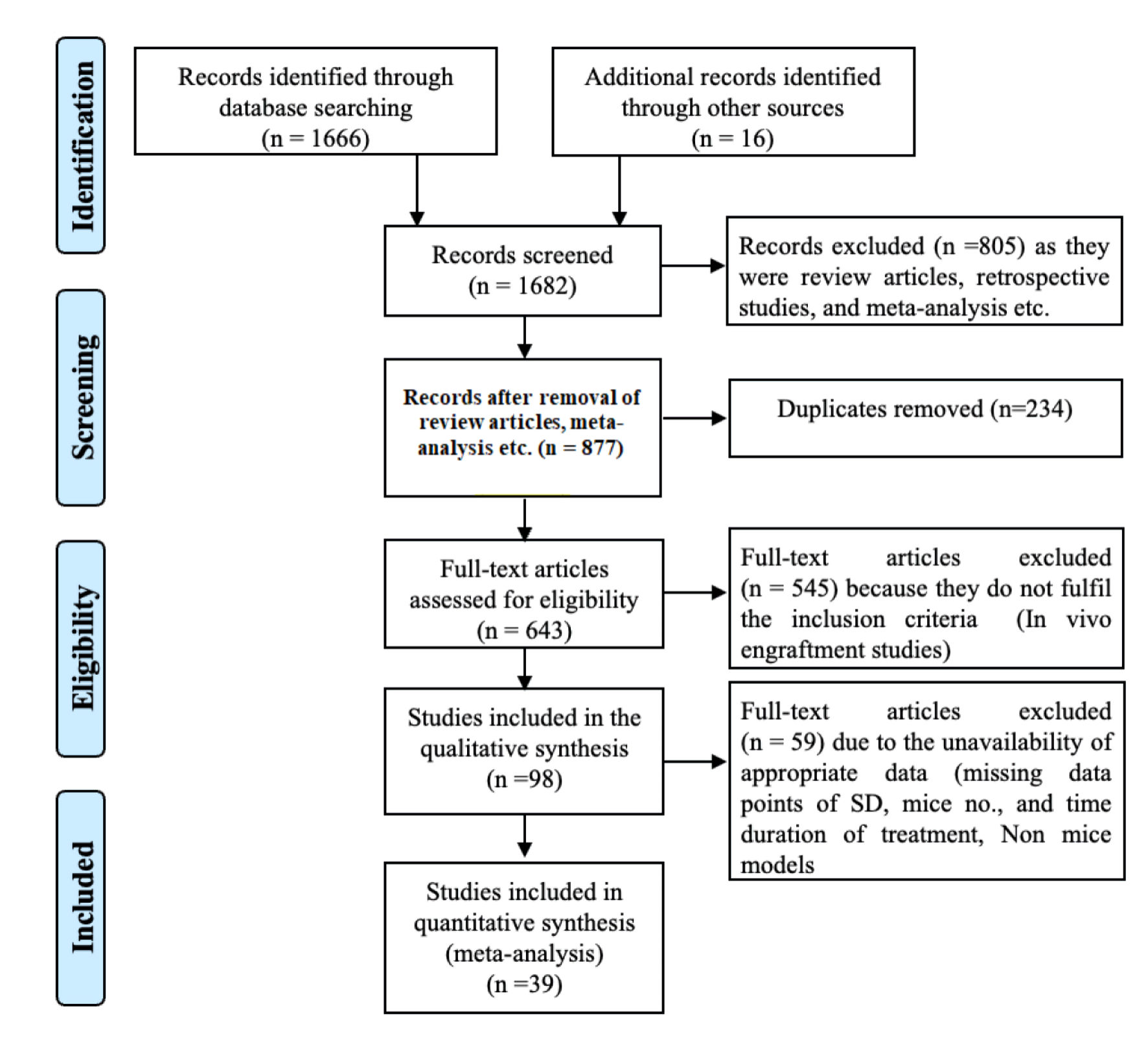 Click for large image | Figure 1. Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) flowchart for the systematic literature search and study inclusion. |
Data extraction
Three investigators (SY, BC, and BA) independently screened the literature, resolving any disagreements through discussions with additional authors (AK, SY, and PKS). Data were quantitatively extracted from bar plots in each article using the WebPlotDigitizer program and from the tables, subsequently organized in a Microsoft Excel spreadsheet (Windows 10 edition; Microsoft Corporation, Lisbon, Portugal) to record species and strains, the number of animals/groups, model cell lines, route of administration of cell used for engraftment in animal models.
Quantitative data analysis
Pooled data analysis was conducted using Jamovi Software Version 2.6.45.0, with Hedge’s g selected as the “effect size” metric [18]. Heterogeneity among studies was evaluated using Cochran’s Q test and the heterogeneity index (I2). A significance threshold of P < 0.10 was applied because of the sensitivity of the test. The quantitative assessment of heterogeneity was based on the I2 scale: low (< 25%), moderate (50–75%), and high (> 75%). The fixed or random effects model was chosen to compute the pooled effect size based on the level of heterogeneity. When the I2 value was ≤ 50%, fixed effect models were applied; however, data with an I2 value > 50% were analyzed using the random effect model. Subgroup analysis was conducted based on mouse strains, knock-in or knockout strategy, mode of transplantation, source of HSPC, CRISPR system, homology-directed repair (HDR) template, and targeted disease.
Sensitivity analysis
Sensitivity analysis was conducted by systematically excluding one study at a time to assess its impact on the pooled effect size estimate. This approach was used to evaluate the influence of individual studies on overall findings.
Publication bias analysis
We conducted an analysis of publication bias by qualitatively examining the asymmetry in the funnel plot and quantitatively based on Egger’s intercept and Begg and Mazumdar rank test.
Institutional review board approval and ethical compliance with human or animal studies were not applicable for this study.
| Results | ▴Top |
Study design and parameters measured
A comprehensive literature search yielded 1,682 potential articles, of which 39 studies were included. These studies targeted 18 different gene loci, distributed as follows: HBB (n = 8), HBG (n = 5), CD33 (n = 4), CCR5, ELANE, and BCL11 (n = 3 each), CYBB, CD45 (n = 2 each), and TET2, Cohesin, ITGB2, CD40L, FXN, FOXP3, MAGT1, PKLR, WAS, IL2RG, and BTK (n = 1 each). All these studies employed Cas9 as a nuclease delivered in the form of ribonucleoprotein (ribonucleoprotein complex (RNP), n = 31), RNA (n = 4), plasmid (n = 3), and virus (n = 1) via electroporation (n = 36), transfection (n = 2), and nanoparticles and transduction (n = 1 each). These articles utilized Cas9 for genomic modifications through knockout and knock-in (n = 19 each), while one study performed base editing. During gene editing via knock-in, HDR templates were delivered as single-strand oligonucleotide (ssODNs, n = 6) and through adeno-associated virus (AAV, n = 15), where two studies used both, as indicated in Table 1 [19–57]. The literature review and study screening results are shown in the PRISMA flow diagram in Figure 1.
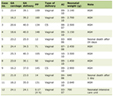 Click to view | Table 1. Characteristic Summary of the Preclinical Studies Included in the Metanalysis |
The majority of the studies utilized mobilized PB stem cells (n = 32), while some used UCB stem cells and BM-derived HSPCs (n = 3 each). One study utilized fetal liver derived stem cells. Mobilized PB HSPCs and BM-derived HSPCs were isolated from patient samples or healthy individuals. These cells were purified using antibodies against the CD34 epitope. Purified CD34+ cells were suspended in 1 × phosphate-buffered saline (PBS) or 1% saline at various concentrations for injection into mice. The total number of cells injected per mouse ranged from 0.09 × 106 to 3 × 106 cells. Specifically, 20 studies used 0.09–0.5 × 106 cells, 18 studies used 0.5–1 × 106 cells, seven studies used 1–1.5 × 106 cells, and six studies used 1.5–3 × 106 cells (Table 1) [19–57]. Some studies have employed varying cell quantities across different experiments.
We have provided details of the mice models and transplantation techniques used in all investigations (Table 1) [19–57]. Most of the studies utilized the tail vein as the preferred transplantation route (24 studies). Alternative methods included retro-orbital (six studies), intraperitoneal (one study), intrahepatic (five studies), and intra-femoral (two studies), while one publication did not provide the route of transplantation [38]. All of these studies utilized four different mouse strains, NSG (23 studies), NSGS, NSGSGM3 (three studies each), NBSGW (nine studies), and one study used the NOG-EXL mouse strain. The follow-up period for engraftment analysis varied from 8 weeks to 24 weeks, where most of the experiments were terminated between 12 and 16 weeks (25 studies), while other studies monitored the engraftment within 8–12 weeks (eight studies), 16–20 weeks (seven studies) and 20–24 weeks (four studies), as detailed in Table 1 [19–57].
We measured the engraftment potential of gene edited versus unedited cells in the BM, spleen, thymus and PB (Table 2). Human cell chimerism in all transplanted mice was analyzed by flow cytometry using CD45 as a marker, while two studies used human leukocyte antigen ABC (HLA-ABC) for quantification because they had deleted the CD45 locus for their study.
Click to view | Table 2. Summary of the Pooled Data and Subgroup Analysis of Various Parameters of the Study |
Effect of CRISPR/Cas9-edited HSPCs on engraftment potential in BM, PB, spleen, and thymus
In a pooled group analysis following the fixed effect model, a significant difference was observed in the mean difference between gene-edited and unedited cells. Twenty-nine studies using BM for their engraftment analysis with 68 datasets were incorporated into the pooled analysis (Supplementary Material 1, jh.elmerpub.com). The fixed effect model analysis demonstrated a significant reduction in the engraftment of gene-edited cells in the BM (95% confidence interval (CI), –0.292 to –0.028; P = 0.018) (Fig. 2a). The findings of the studies were unbiased, as neither the rank correlation nor the regression test indicated any funnel plot asymmetry (P = 0.1373 and P = 0.7823, respectively) (Supplementary Material 2A, jh.elmerpub.com). Additionally, 11 studies presented 24 datasets for spleen engraftment (Supplementary Material 3, jh.elmerpub.com), and 24 studies presented 41 datasets for PB engraftment (Supplementary Material 4, jh.elmerpub.com). The fixed effect model analysis revealed a significant decline in the engraftment of gene-edited HSPCs in the spleen and PB (95% CI, –0.542 to –0.097; P = 0.005; 95% CI, –0.464 to –0.075; P = 0.007, respectively) (Fig. 2b, c). However, the spleen data were homogenous in nature (Tau2 = 0.0316; I2 = 9.07%) and biased, as both the rank correlation and regression tests indicated potential funnel plot asymmetry (P < 0.0001 and P = 0.0016, respectively) (Supplementary Material 2B, jh.elmerpub.com). Conversely, the PB data were homogeneous and unbiased, as evidenced by funnel plot symmetry (correlation P = 0.9023 and regression P = 0.7030) (Supplementary Material 2C, jh.elmerpub.com). Furthermore, the data presented for thymus engraftment (five studies, 11 datasets) indicate no discernible difference in engraftment capacity between gene-edited and unedited HSPCs (Supplementary Material 5, jh.elmerpub.com).
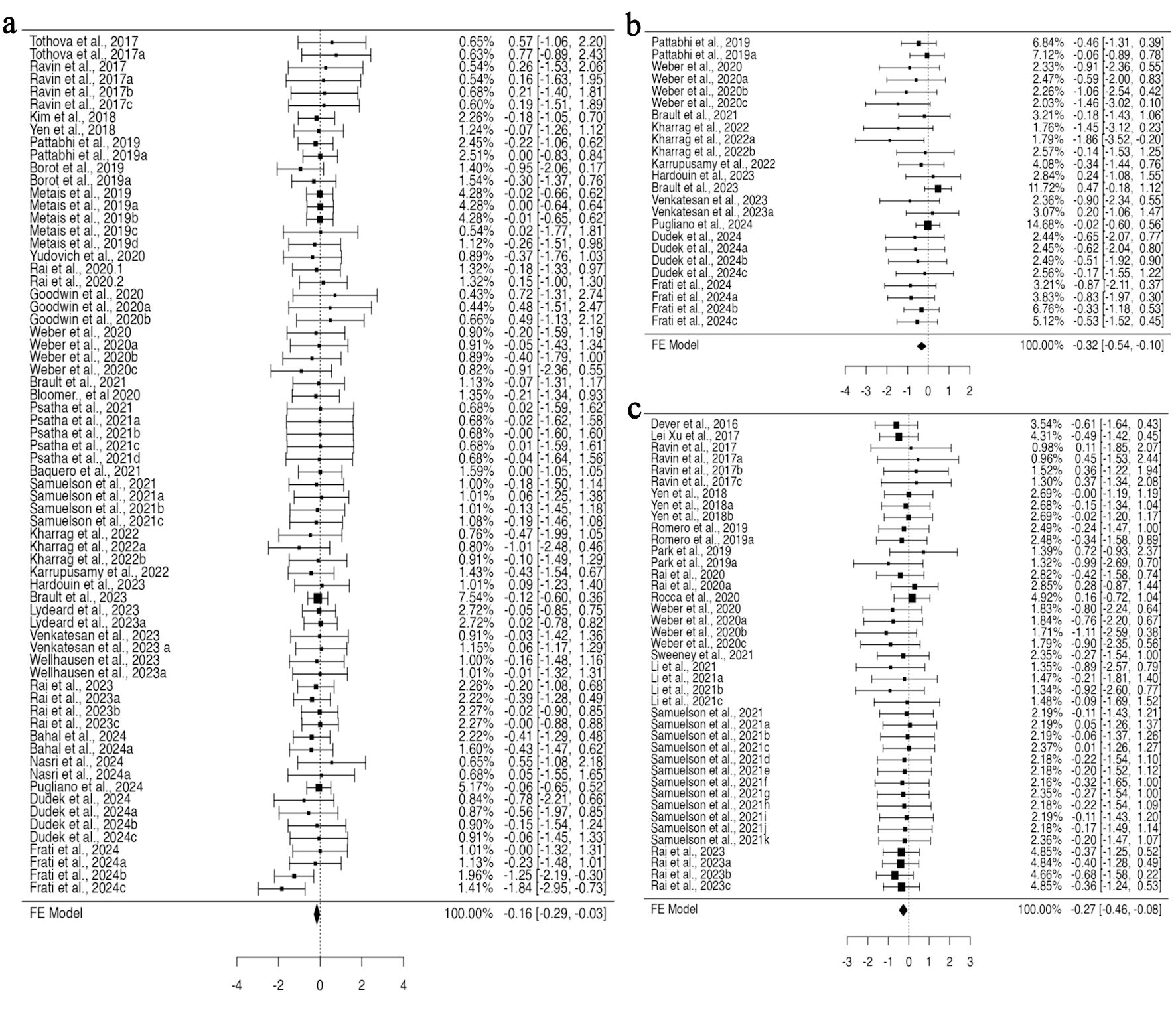 Click for large image | Figure 2. CRISPR-Cas9 gene editing negatively impacts the ability of hematopoietic stem and progenitor cells to engraft in vivo, as measured by fluorescent-activated cell sorting. (a) Analysis of bone marrow engraftment comprised a total of 68 datasets. Most estimates were negative (68%), with observed standardized mean differences ranging from –1.8391 to 0.7740. (b) Analysis of spleen engraftment comprised a total of 24 datasets. Most estimates were negative (88%), and the observed standardized mean differences varied from –1.8629 to 0.4730. (c) Analysis of peripheral blood engraftment comprised a total of 41 datasets. The majority of estimates (78%) were negative, with observed standardized mean differences ranging from –1.1051 to 0.7224. CRISPR-Cas9: clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9. |
Effect of CRSIPR/Cas9 gene editing strategy through knock-in or knockout on engraftment potential
Gene editing encompasses both the insertion of DNA (knock-in) and the deletion of genes (knockout), and both methodologies were reported in the included studies. HSPCs genetically modified through knock-in did not exhibit a significant difference in the standardized mean difference compared to non-edited cells for the data provided across all analyzed organs. Conversely, knockout HSPCs demonstrate significant variations in engraftment efficacy in specific organs. For BM engraftment, 13 studies provided 28 datasets for knock-in, and 15 studies provided 38 datasets for knockout HSPCs. A fixed effect model applied to knockout HSPCs revealed a statistically significant standardized mean difference (95% CI, –0.401 to –0.035; P = 0.020) (Fig. 3a), suggesting a decrease in the engraftment of knockout HSPCs in the BM; however, the data were homogenous without any funnel plot asymmetry (Supplementary Material 6A, jh.elmerpub.com). For PB engraftment, 11 studies provided 33 datasets for the knockout, and 34 datasets from 13 studies for knock-in HSPCs. There was no heterogeneity; however, some bias in the data was determined based on Tau2 and funnel plot asymmetry (Supplementary Material 6C, jh.elmerpub.com). Using a fixed effect model, the data analysis revealed a significant difference in the standardized mean difference measure (95% CI, –0.442 to –0.020; P = 0.032) (Fig. 3c). This concedes the lower engraftment of knockout HSPCs in the PB compared to unedited counterparts. Among the 12 articles providing data on spleen engraftment, 14 experimental studies contributed data on gene knockout and nine on knock-in. The available data exhibited no heterogeneity but were slightly biased, as revealed by the Tau2 and funnel plot asymmetry (Supplementary Material 6B, jh.elmerpub.com). The fixed effect model analysis concedes a highly significant standardized mean difference for the engraftment of knockout HSPCs in the spleen compared to non-edited ones (95% CI, –1.00 to –0.327; P < 0.001) (Fig. 3b).
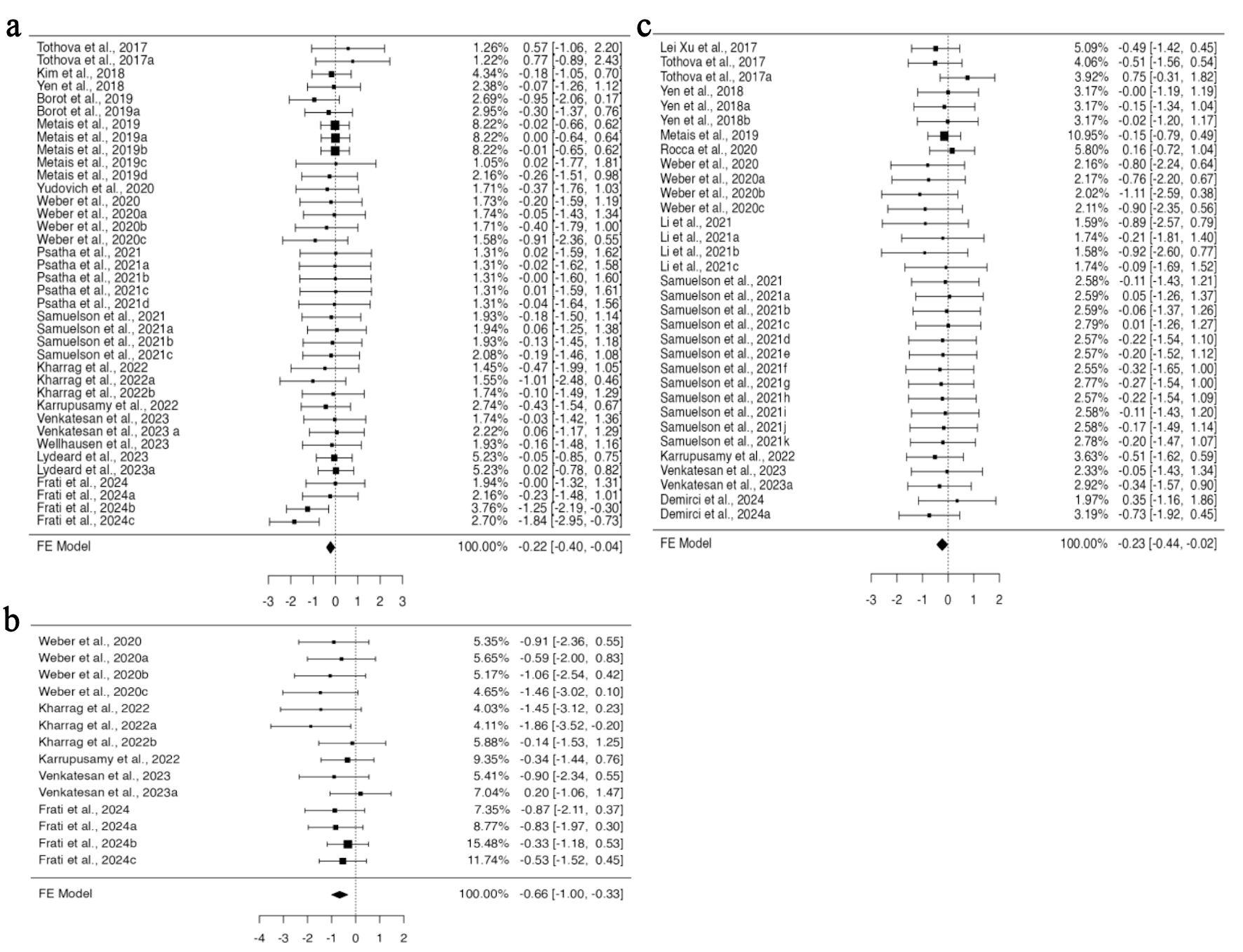 Click for large image | Figure 3. The in vivo engraftment potential of human CD34+ HSPCs is impacted by the gene knockout approach used for gene editing. (a) Analysis of bone marrow engraftment included 38 datasets. The majority of estimates (76%) were negative, with observed standardized mean differences ranging from –1.8391 to 0.7740. (b) Analysis of spleen engraftment comprised a total of 14 datasets, of which 93% of the estimates were negative, with the observed standardized mean differences ranging from –1.8629 to 0.2046. (c) Analysis of peripheral blood engraftment comprised a total of 33 datasets. Most estimates (85%) were negative, with the observed standardized mean differences ranging from –1.1051 to 0.7534. HSPCs: hematopoietic stem and progenitor cells. |
The consequences of CRISPR-Cas9 system used for gene editing on the engraftment of gene-edited HSPCs
To perform gene editing, guide RNA and CRISPR-Cas9 can be introduced into cells as RNA, RNP, or in the form of plasmids. Among all the studies included in the BM engraftment analysis, 25 articles provided 61 investigations with data for RNP, while seven datasets for RNA from four studies, for gRNA/CRISPR-Cas9 delivery system. No heterogeneity was found in the data for RNP (Supplementary Material 7A, jh.elmerpub.com); therefore, the fixed effect model analysis suggested a significant variation in the standardized mean difference (95% CI, –0.318 to –0.037; P = 0.014) (Fig. 4a). This signifies decreased engraftment of gene-edited HSPCs in the BM when using RNP as a delivery system for CRISPR-Cas9, while no effect was observed when using them in RNA form. For the investigation of spleen engraftment of gene-edited HSPCs, eight studies from 21 experiments provided the data for RNP, and three studies provided the three experimental data for RNA. Using the fixed effect model, the standardized mean difference yielded a highly significant value for RNP-mediated delivery (95% CI, –0.702 to –0.212; P < 0.001) (Fig. 4c). The analysis revealed altered engraftment of RNP-mediated gene-edited HSPCs, whereas no effect was observed when RNA was used. However, the available data showed a potential funnel plot asymmetry, suggesting a bias in the study (Supplementary Material 7B, jh.elmerpub.com). In total, for the PB engraftment study, 21 studies with 60 experiments provided the data for RNP and eight for RNA from three studies. A fixed effect model was selected to measure the outcome of the standardized mean difference as the data were homogenous and unbiased (Supplementary Material 7C, D, jh.elmerpub.com). The outcome showed the decreased engraftment of RNP and increased engraftment of RNA-mediated gene-edited HSPCs in the spleen (95% CI, –0.388 to –0.096; P = 0.001; 95% CI, 0.046 to 0.649; P = 0.024, respectively) (Fig. 4b, d).
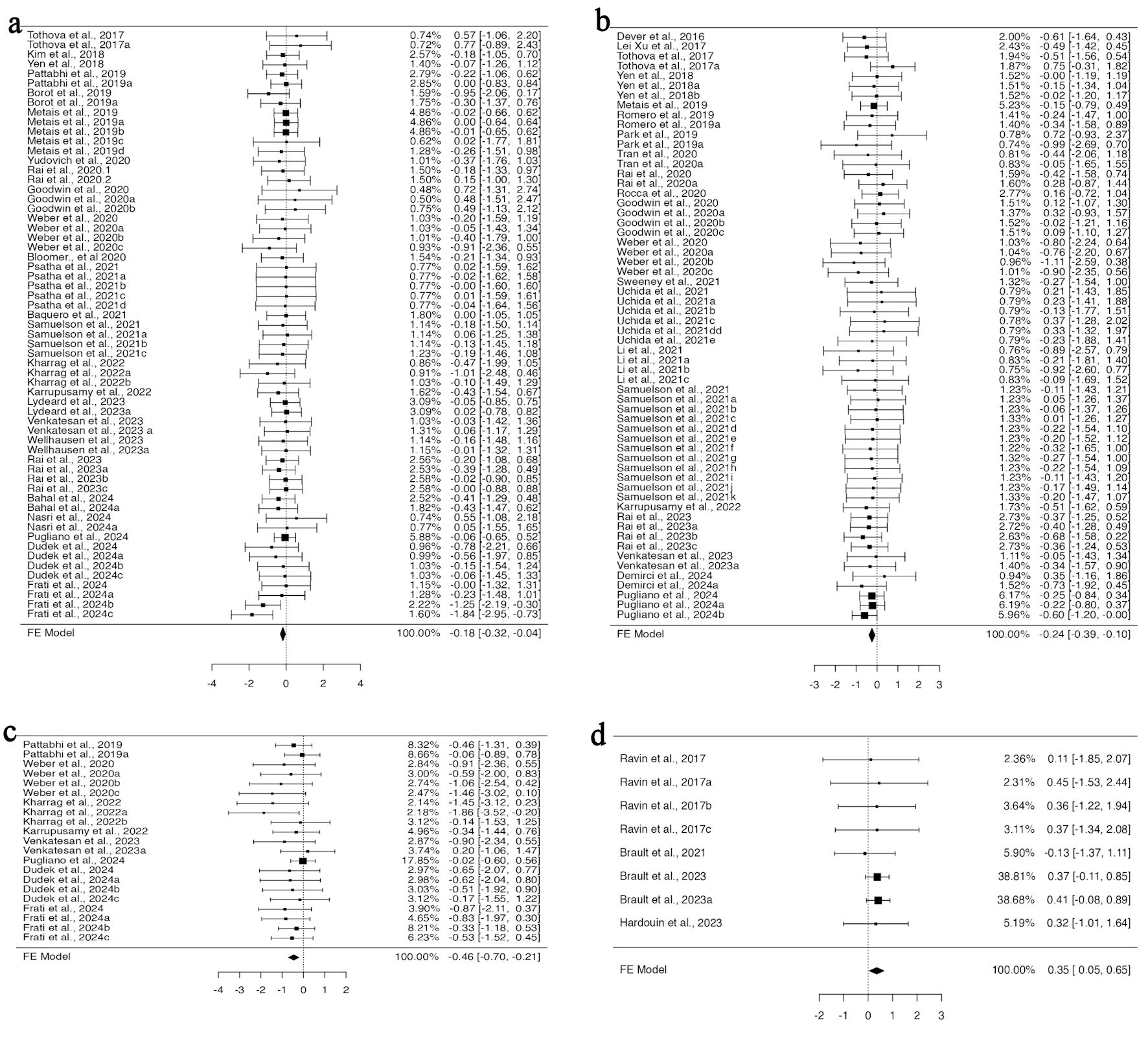 Click for large image | Figure 4. CRISPR-Cas9-mediated gene editing using RNP complex affects the in vivo engraftment of human HSPCs in mice model. (a) The analysis of bone marrow engraftment comprised 61 datasets that used the RNP version of the CRISPR-Cas9 system. The majority of estimates (72%), with observed standardized mean differences ranging from –1.8391 to 0.7740, were negative. (b) The study of peripheral blood engraftment included 60 datasets that used the RNP version of the CRISPR-Cas9 technology. A significant proportion of estimates (77%), with observed standardized mean differences ranging from –1.1051 to 0.7534, were negative. (c) Twenty-one datasets in all that used the RNP version of the CRISPR-Cas9 technology were analyzed for spleen engraftment. The observed standardized mean differences were mostly negative (95%), with a range of –1.8629 to 0.2046. (d)The investigation of peripheral blood engraftment comprised a total of eight datasets that used the CRISPR-Cas9 system in RNA form. The majority of estimates (88%), with observed standardized mean differences ranging from –0.1343 to 0.4505, were positive. CRISPR-Cas9: clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9; HSPCs: hematopoietic stem and progenitor cells; RNP: ribonucleoprotein complex. |
The implications of mouse strain used in engraftment studies for CRISPR-Cas9-mediated gene-edited HSPCs
In the 39 publications included in our study, four distinct mouse strains were used. When employing NBSGW and NSGSGM3 mouse strains, subgroup analysis based on mouse strain using a fixed effect model revealed no difference in the engraftment of edited cells compared to non-edited HSPCs in all four organs investigated. However, when the NSGS mouse model was used for HSPC transplantation, there was a significant increase in the engraftment of HSPCs in the PB (95% CI, 0.011 to 0.615; P = 0.042) (Fig. 5c). The data analyzed using a fixed effect model from five datasets across two studies were homogenous, with no bias inferred from funnel plot asymmetry (Supplementary Material 8C, jh.elmerpub.com). Additional subgroup analysis using the NSG mouse model, based on 38 datasets from 29 studies for BM and 11 studies with 11 datasets for spleen engraftment, indicated a significant reduction in the engraftment of gene-edited cells (95% CI, –0.426 to –0.051; P = 0.013; 95% CI, –1.145 to –0.383; P < 0.001, respectively) (Fig. 5a, b). Neither the rank correlation nor the regression test indicated any funnel plot asymmetry in the data provided for the BM; however, the data for spleen showed some bias (Supplementary Material 8A, B, jh.elmerpub.com).
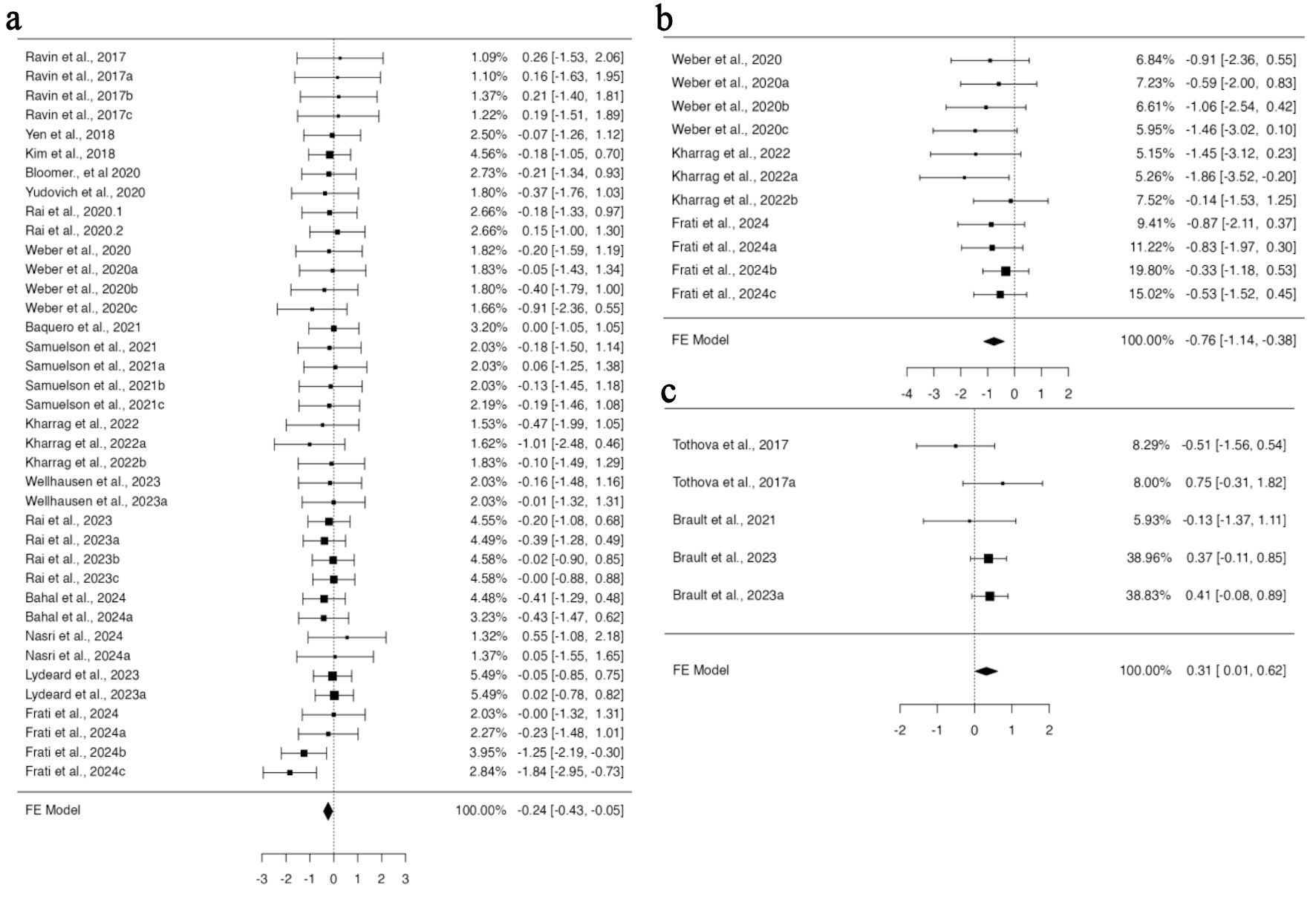 Click for large image | Figure 5. The genetic background of NSGS and NSG Inbred mice influences the engraftment of gene-edited hHSPCs in the spleen, bone marrow, and peripheral blood. (a) The analysis of bone marrow engraftment comprised 38 datasets that used the NSG strain of mice model. The majority of estimates (74%), with observed standardized mean differences ranging from –1.8391 to 0.5537, were negative. (b) Analysis of spleen engraftment was conducted on 11 datasets that used the NSG strain of mice model. All estimations were negative (100%), and the observed standardized mean differences varied from –1.8629 to -0.1399. (c) The analysis comprised five datasets on peripheral blood engraftment using the NSGS mouse strain model. The observed standardized mean differences varied from –0.5092 to 0.7534, with 60% of the estimations being positive. hHSPCs: human hematopoietic stem and progenitor cells. |
Impact of different cell delivery methods for CRISPR-Cas9-mediated gene-edited HSPCs on organ-specific engraftment
We categorized the studies into subgroups based on the different routes of cell administration. Five distinct routes of cell administration were identified across all 39 studies. In all, 11 experiments from four studies provided data for BM engraftment through the retro-orbital route of transplantation, whose funnel plot was asymmetrical for the regression test but not for the correlation one (Supplementary Material 9A, jh.elmerpub.com). Fixed model analysis revealed deficient engraftment of gene-edited cells compared to unedited ones (95% CI, –0.894 to –0.109; P = 0.012) (Fig. 6a). For spleen engraftment, eight datasets from four studies utilized tail vein injection, 10 datasets from four studies utilized the retro-orbital route, and four datasets from one study preferred the intraperitoneal route. Data provided in all subgroups were independently analyzed using a fixed effect model. There was no heterogeneity or bias in the data of any subgroup (Supplementary Material 9B–D, jh.elmerpub.com); however, the engraftment potential of gene-edited cells was found to be minimized. The standardized mean difference was statistically significant in the tail vein (95% CI, –0.837 to –0.019; P = 0.04) (Fig. 6b), retro-orbital (95% CI, –0.657 to –0.004; P = 0.047) (Fig. 6c), and intra-peritoneal (95% CI, –1.721 to –0.245; P = 0.009) (Fig. 6d) subgroups.
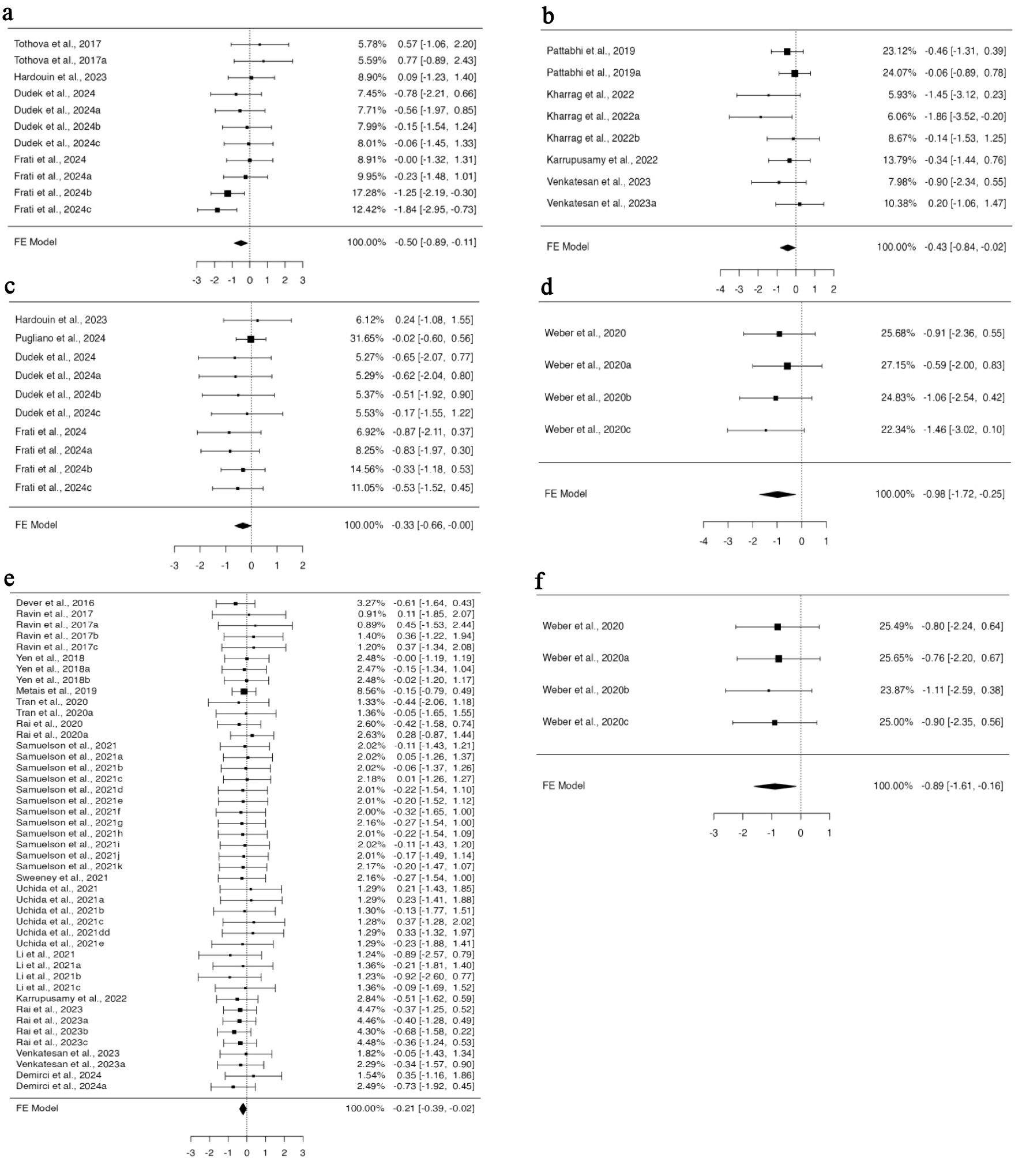 Click for large image | Figure 6. Utilizing the tail vein, intraperitoneal, and retro-orbital routes to deliver cells compromises the engraftment efficiency of gene-edited hHSPCs in the spleen and peripheral blood. The analysis of spleen engraftment comprised (a) eight datasets on spleen engraftment that used the tail vein as the delivery route. The observed standardized mean differences varied from –1.8629 to 0.2046, with 88% of estimates being negative. (b) Ten datasets that used retro-orbital administration. The observed standardized mean differences varied from –0.8680 to 0.2357, with 90% of the estimations being negative. (c) A total of four datasets that used intraperitoneal as the delivery method. All estimations were negative (100%), and the observed standardized mean differences varied from –1.4638 to -0.5878. In order to analyze peripheral blood engraftment (d), 45 datasets that used tail vein injection were considered. A significant portion of estimates (73%), with observed standardized mean differences ranging from –0.9161 to 0.4505, were negative. (e) A total of four datasets with intraperitoneal delivery were included. All estimations were negative (100%), and the observed standardized mean differences varied from –1.1051 to –0.7621. hHSPCs: human hematopoietic stem and progenitor cells. |
In total, 14 studies provided 45 datasets on PB engraftment via tail vein transplantation, nine datasets from four studies for intrahepatic, eight datasets from four articles for retro-orbital, four datasets for intraperitoneal, and two datasets for intra-femoral from one study each. The cells transplanted through the tail vein and intraperitoneal route engrafted less efficiently when edited with CRISPR-Cas9 compared to non-edited ones. A fixed effect model was fitted to the analysis, which yielded a statistically significant value for the standardized mean differences (95% CI, –0.394 to –0.020; P = 0.030) for the tail vein (Fig. 6e) and (95% CI, –1.614 to –0.160; P = 0.017) for the intraperitoneal (Fig. 6f) routes. No heterogeneity was found in the data for either subgroup. Neither the rank correlation nor the regression test indicated any funnel plot asymmetry for the data in the intraperitoneal subgroup (P = 0.0833 and P = 0.7220, respectively), while the rank correlation test indicated funnel plot asymmetry (P = 0.0127) for the tail vein subgroup, indicating bias in the data (Supplementary Material 9E, F, jh.elmerpub.com).
The influence of several tissue-specific HSPCs obtained from various sources on cell engraftment after gene editing
There are four sources of HSPCs: BM, PB, UCB, and fetal liver, from which cells are obtained for gene editing and transplantation. After analysis using a fixed effect model, HSPCs from PB showed reduced engraftment in the BM and PB. In BM engraftment studies, 58 investigations from 22 publications used HSPCs from PB, and six datasets each from BM and UCB from three and four publications, respectively. The analysis was carried out using the standardized mean difference as the outcome measure, using a fixed effect model that showed a significant difference in the outcome for HSPCs from PB (95% CI, –0.305 to –0.027; P = 0.019) (Fig. 7a). Of the 25 studies that provided data on PB engraftment, 23 publications with 60 datasets used PB-derived HSPCs, while one each with four datasets used BM- and UCB-derived HSPCs. The estimated average standardized mean difference based on the fixed effects model differed significantly (95% CI, –0.324 to –0.035; P = 0.015), suggesting decreased engraftment of PB-derived gene-edited HSPCs in PB (Fig. 7b). There was no heterogeneity in the data, nor was there any funnel plot asymmetry (Supplementary Material 10A, B, jh.elmerpub.com).
 Click for large image | Figure 7. The engraftment capacity of cells is affected by CRISPR-Cas9-based gene editing of peripheral blood-derived HSPCs and hemoglobinopathy-related genes. The investigation of HSPCs obtained from peripheral blood in (a) bone marrow engraftment included 58 datasets in total. With 72% of estimations being negative, the observed standardized mean differences varied from –1.8391 to 0.7740. (b) Peripheral blood engraftment included a total of 60 datasets. The observed standardized mean differences ranged from –1.1051 to 0.7534, with 70% of estimations being negative. To examine the editing effect of hemoglobinopathy-related genes (c) on peripheral blood engraftment, 40 datasets were analyzed. The majority of estimates (78%), with observed standardized mean differences ranging from –1.1051 to 0.7224, were negative. (d) For spleen engraftment, data from 13 datasets were utilized. Most estimates (85%) were negative, and the observed standardized mean differences varied from –1.4431 to 0.2338. CRISPR-Cas9: clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9; HSPCs: hematopoietic stem and progenitor cells. |
The impact of disease-specific gene editing using the CRISPR-Cas9 approaches on HSPC engraftment efficiency
A total of 18 gene loci were targeted across the 39 studies included in this analysis, which were associated with five distinct diseases: human immunodeficiency virus (CCR5), immune-mediated disease (CYBB, ITGB2, FOXP3, MAGT1, WAS, BTK, CD40L), hemoglobinopathies (HBB, HBG, BCL11), leukemia (TET2, cohesin, CD33, CD45), and severe congenital neutropenia (ELANE). Transplantation of gene-edited HSPCs in various disease conditions improved the overall outcomes in mouse models. The engraftment of CRISPR-Cas9-mediated genetically modified HSPCs was found to be comparable to that of unedited HSPCs in all disease conditions analyzed in the BM and thymus. In the spleen and PB, except for hemoglobinopathy, gene-edited HSPCs for other targeted disease conditions engrafted similarly to the control. Homogeneous and unbiased data on PB engraftment of HSPCs for CRISPR-Cas9 based modifications of genetic loci involved in hemoglobinopathy were provided by 12 studies with 40 datasets (Supplementary Material 11C, jh.elmerpub.com). The engraftment of gene-edited HSPCs was found to be diminished when analyzed using the fixed effect model (95% CI, –0.432 to –0.016; P = 0.035) (Fig. 7c).
A total of five studies provided 13 datasets for spleen engraftment targeting hemoglobinopathy, which were biased (Supplementary Material 11A, jh.elmerpub.com), and three datasets from one study targeted leukemia genes, which were homogenous and unbiased (Supplementary Material 11B, jh.elmerpub.com). A fixed effect model analysis revealed less engraftment of gene-edited cells targeted for hemoglobinopathy compared to non-edited cells (95% CI, –0.793 to –0.166; P = 0.003) (Fig. 7d) and leukemia genes (95% CI, –1.919 to –0.121; P = 0.026) (Fig. 7e).
| Discussion | ▴Top |
Allogeneic HSPC therapy for hereditary blood disorders is limited by donor availability and immunological complications, whereas autologous HSPC gene therapy offers a safer alternative. Recent advances in vector-mediated gene addition and CRISPR-Cas9 gene editing have expanded the therapeutic potential of genetic disorders. Preclinical studies remain crucial for assessing the engraftment efficiency of edited HSPCs in organs such as the BM, spleen, thymus, and PB. Our systematic review and meta-analysis showed that gene-edited cells exhibit lower engraftment efficiency than unmodified cells, except in the thymus. Subgroup analysis revealed that knock-in cells were engrafted similarly to unaltered cells, whereas knockout cells showed reduced engraftment, reflecting the challenges of HDR, which requires the co-delivery of a donor template (ssODN or AAV6). No significant difference in engraftment was observed between the HDR templates, suggesting that the template type may not influence engraftment outcomes. Enhancing HSPC proliferation may partially mitigate reduced engraftment but often compromises stemness, emphasizing the need for improved gene editing techniques (Table 3).
Click to view | Table 3. Practical Recommendations for Optimizing Engraftment |
CRISPR-Cas9 enables precise and regulated gene correction, avoiding the genotoxic risks associated with semi-random viral vector integration. Nevertheless, the success of this platform is contingent upon the meticulous selection of reagents to achieve a balance between efficacy, specificity, and safety while minimizing cytotoxicity, especially in self-renewing HSPCs. Our subgroup analysis revealed that the CRISPR-Cas9 delivery mode significantly influenced engraftment outcomes. Specifically, RNP-based delivery reduced engraftment in both the BM and spleen, whereas both RNP and RNA delivery decreased PB engraftment. Although plasmid-based CRISPR-Cas9 systems are simpler and prevent multi-component transfection, and Cas9 mRNA systems exhibit reduced stability, the limited data available preclude robust statistical conclusions. Crucially, further investigation is required to assess the genome integrity of edited HSPCs, as emerging evidence indicates the potential risks of chromosomal arm loss, translocations, deletions, and chromothripsis following nuclease treatment [58–60].
Engraftment outcomes are influenced by the HSPC source. Each source BM, PB, or cord blood has distinct advantages regarding engraftment kinetics, immune recovery, and GVHD risk. Preclinical studies have relied on animal models to explore HSPC transplantation and gene editing. Repeated HSPC transplantation leads to exhaustion, although primary transplants achieve robust engraftment across organs [61]. Our findings showed that the administration route and animal strain affected engraftment potential. Spleen engraftment diminished when cells were delivered via the tail vein, intraperitoneal, or retro-orbital routes, whereas PB engraftment was compromised following tail vein or intraperitoneal injection. The spleen, a potential extramedullary hematopoietic site, contributes to early cytokine-driven engraftment reactions. Splenectomy facilitates early engraftment, whereas splenomegaly delays it, possibly explaining the diminished splenic engraftment [62].
Engraftment differences across mouse strains reflect immune modulation. NSG and NSGS mice, which lack T, B, and natural killer (NK) cells, showed lower engraftment of gene-edited HSPCs than NBSGW and NSG-SGM3 strains, which possess modifications facilitating enhanced human HSPC engraftment [63]. While pooled analyses indicated reduced persistence of gene-edited HSPCs in the BM, disease-specific subgroups showed no consistent impact on transplantation efficiency. In HIV, immune-mediated disorders, hemoglobinopathies, leukemia, and severe congenital neutropenia models, CRISPR-Cas9–corrected HSPCs maintained robust engraftment. Gene-edited HSPCs restored immune function and conferred HIV-1 resistance; corrected hemoglobinopathy models showed reduced HbS levels, mimicking asymptomatic phenotypes; and edited HSPCs improved outcomes in immune-mediated disease and neutropenia models. In chimeric antigen receptor T-cells (CAR-T) modifications, HSPC gene editing reduces tumor burden while preserving stem cell function. CRISPR-Cas9–based HSPC gene therapy shows promise but faces challenges in optimizing delivery, preserving stemness, and ensuring genomic stability. Understanding the effects of editing on engraftment will be crucial for translating preclinical success into clinical therapies.
Limitations
The nonhuman primate (NHP) autologous transplantation model is a valuable tool for assessing the long-term durability of gene-modified cells; however, it was excluded because it does not use human hematopoietic cells. Our analysis is subject to several limitations, including the lack of reported blinding and randomization, unaccounted ex vivo HSPC culture conditions and cell doses, and uneven study weighting resulting from variance differences. These issues highlight the necessity of standardized protocols in transplantation, cell culture, and gene editing, as well as preregistered study designs. The standardization of these procedures would facilitate more comprehensive information synthesis initiatives, which should hasten the transition to clinical trials.
| Supplementary Material | ▴Top |
Suppl 1. Assessment of engraftment percentage in bone marrow for CRISPR-Cas9-edited HSPCs.
Suppl 2. Funnel plot for pooled analysis.
Suppl 3. Comprehensive evaluation of CRISPR-Cas9 gene-edited HSPC engraftment in spleen.
Suppl 4. Evaluation of the percentage of CRISPR-Cas9 gene-edited HSPCs that engraft in peripheral blood.
Suppl 5. Comprehensive measurement of engraftment for CRISPR-Cas9 gene-edited HSPCs in thymus.
Suppl 6. Funnel plot of knockout subgroup analysis.
Suppl 7. Funnel plot of subgroup for CRISPR system analysis.
Suppl 8. Funnel plot of mice strain subgroup analysis.
Suppl 9. Funnel plot for subgroup route of delivery analysis.
Suppl 10. Funnel plot for the subgroup analysis of peripheral blood as source of HSPC analysis.
Suppl 11. Funnel plot for subgroup analysis of disease-targeted and HDR analyses.
Acknowledgments
This research was supported by the Intramural Research Grant from Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow.
Financial Disclosure
This research was funded by the Intramural Research Grant from Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow to Sudhanshu Yadav.
Conflict of Interest
Author, Shubhanshu Yadav is a professional receiving financial support from IQVIA. The remaining authors declare no competing financial interests.
Informed Consent
Not applicable.
Author Contributions
SY designed research, screened paper, curated and analyzed data, acquired funding and wrote the paper. BC and BA designed research, screened paper, analyzed data and wrote the paper. SY, PKS, AK searched literature, reviewed and edited the paper.
Data Availability
The authors declare that data supporting the findings of this study are available within the article.
| References | ▴Top |
- Barriga F, Ramirez P, Wietstruck A, Rojas N. Hematopoietic stem cell transplantation: clinical use and perspectives. Biol Res. 2012;45(3):307-316.
doi pubmed - Eapen M, Brazauskas R, Walters MC, Bernaudin F, Bo-Subait K, Fitzhugh CD, Hankins JS, et al. Effect of donor type and conditioning regimen intensity on allogeneic transplantation outcomes in patients with sickle cell disease: a retrospective multicentre, cohort study. Lancet Haematol. 2019;6(11):e585-e596.
doi pubmed - Niederwieser D, Baldomero H, Szer J, Gratwohl M, Aljurf M, Atsuta Y, Bouzas LF, et al. Hematopoietic stem cell transplantation activity worldwide in 2012 and a SWOT analysis of the Worldwide Network for Blood and Marrow Transplantation Group including the global survey. Bone Marrow Transplant. 2016;51(6):778-785.
doi pubmed - Imahashi N, Kurita N, Konuma T, Takahashi S, Nishida T, Tanaka M, Nakamae H, et al. Effect of conditioning regimens and graft-versus-host disease prophylaxis on the outcomes of umbilical cord blood transplantation performed with cyclophosphamide/total body irradiation-based regimens. Transplant Cell Ther. 2024;30(3):318.e1-e11.
doi pubmed - John T, Czechowicz A. Clinical hematopoietic stem cell-based gene therapy. Mol Ther. 2025;33(6):2663-2678.
doi pubmed - Fischer A, Hacein-Bey-Abina S. Gene therapy for severe combined immunodeficiencies and beyond. J Exp Med. 2020;217(2):e20190607.
doi pubmed - Gaj T, Gersbach CA, Barbas CF, 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013;31(7):397-405.
doi pubmed - Cavazza A, Santilli G. The future of ex vivo hematopoietic stem cell gene editing: what's next. Regen Med. 2025;20(2-3):73-76.
doi pubmed - Dimitrievska M, Bansal D, Vitale M, Strouboulis J, Miccio A, Nicolaides KH, El Hoss S, et al. Revolutionising healing: gene Editing's breakthrough against sickle cell disease. Blood Rev. 2024;65:101185.
doi pubmed - Bernardo ME, Aiuti A. The role of conditioning in hematopoietic stem-cell gene therapy. Hum Gene Ther. 2016;27(10):741-748.
doi pubmed - Panch SR, Szymanski J, Savani BN, Stroncek DF. Sources of hematopoietic stem and progenitor cells and methods to optimize yields for clinical cell therapy. Biol Blood Marrow Transplant. 2017;23(8):1241-1249.
doi pubmed - Chira S, Jackson CS, Oprea I, Ozturk F, Pepper MS, Diaconu I, Braicu C, et al. Progresses towards safe and efficient gene therapy vectors. Oncotarget. 2015;6(31):30675-30703.
doi pubmed - Geng G, Xu Y, Hu Z, Wang H, Chen X, Yuan W, Shu Y. Viral and non-viral vectors in gene therapy: current state and clinical perspectives. EBioMedicine. 2025;118:105834.
doi pubmed - Moaveni AK, Amiri M, Shademan B, Farhadi A, Behroozi J, Nourazarian A. Advances and challenges in gene therapy strategies for pediatric cancer: a comprehensive update. Front Mol Biosci. 2024;11:1382190.
doi pubmed - Soufizadeh P, Mansouri V, Ahmadbeigi N. A review of animal models utilized in preclinical studies of approved gene therapy products: trends and insights. Lab Anim Res. 2024;40(1):17.
doi pubmed - Poletto E, Colella P, Pimentel Vera LN, Khan S, Tomatsu S, Baldo G, Gomez-Ospina N. Improved engraftment and therapeutic efficacy by human genome-edited hematopoietic stem cells with Busulfan-based myeloablation. Mol Ther Methods Clin Dev. 2022;25:392-409.
doi pubmed - Maganti HB, Bailey AJM, Kirkham AM, Shorr R, Pineault N, Allan DS. Persistence of CRISPR/Cas9 gene edited hematopoietic stem cells following transplantation: a systematic review and meta-analysis of preclinical studies. Stem Cells Transl Med. 2021;10(7):996-1007.
doi pubmed - Mohamed SOO, Mohamed AEA, Salih MSK, Salih KSK, Abdelrahman A, Abdelgadir AGA, Ahmedkaroum MGA, et al. Serum lipid profile abnormalities among beta-thalassemia patients: a systematic review and meta-analysis. Lipids Health Dis. 2024;23(1):388.
doi pubmed - Xu L, Yang H, Gao Y, Chen Z, Xie L, Liu Y, Liu Y, et al. CRISPR/Cas9-mediated CCR5 ablation in human hematopoietic stem/progenitor cells confers HIV-1 resistance in vivo. Mol Ther. 2017;25(8):1782-1789.
doi pubmed - Tothova Z, Krill-Burger JM, Popova KD, Landers CC, Sievers QL, Yudovich D, Belizaire R, et al. Multiplex CRISPR/Cas9-based genome editing in human hematopoietic stem cells models clonal hematopoiesis and myeloid neoplasia. Cell Stem Cell. 2017;21(4):547-555.e548.
doi pubmed - De Ravin SS, Li L, Wu X, Choi U, Allen C, Koontz S, Lee J, et al. CRISPR-Cas9 gene repair of hematopoietic stem cells from patients with X-linked chronic granulomatous disease. Sci Transl Med. 2017;9(372).
doi pubmed - Yen J, Fiorino M, Liu Y, Paula S, Clarkson S, Quinn L, Tschantz WR, et al. TRIAMF: a new method for delivery of Cas9 ribonucleoprotein complex to human hematopoietic stem cells. Sci Rep. 2018;8(1):16304.
doi pubmed - Kim MY, Yu KR, Kenderian SS, Ruella M, Chen S, Shin TH, Aljanahi AA, et al. Genetic inactivation of CD33 in hematopoietic stem cells to enable CAR T Cell immunotherapy for acute myeloid leukemia. Cell. 2018;173(6):1439-1453.e1419.
doi pubmed - Pattabhi S, Lotti SN, Berger MP, Singh S, Lux CT, Jacoby K, Lee C, et al. In vivo outcome of homology-directed repair at the HBB gene in HSC using alternative donor template delivery methods. Mol Ther Nucleic Acids. 2019;17:277-288.
doi pubmed - Borot F, Wang H, Ma Y, Jafarov T, Raza A, Ali AM, Mukherjee S. Gene-edited stem cells enable CD33-directed immune therapy for myeloid malignancies. Proc Natl Acad Sci U S A. 2019;116(24):11978-11987.
doi pubmed - Romero Z, Lomova A, Said S, Miggelbrink A, Kuo CY, Campo-Fernandez B, Hoban MD, et al. Editing the sickle cell disease mutation in human hematopoietic stem cells: comparison of endonucleases and homologous donor templates. Mol Ther. 2019;27(8):1389-1406.
doi pubmed - Metais JY, Doerfler PA, Mayuranathan T, Bauer DE, Fowler SC, Hsieh MM, Katta V, et al. Genome editing of HBG1 and HBG2 to induce fetal hemoglobin. Blood Adv. 2019;3(21):3379-3392.
doi pubmed - Bloomer H, Smith RH, Hakami W, Larochelle A. Genome editing in human hematopoietic stem and progenitor cells via CRISPR-Cas9-mediated homology-independent targeted integration. Mol Ther. 2021;29(4):1611-1624.
doi pubmed - Yudovich D, Backstrom A, Schmiderer L, Zemaitis K, Subramaniam A, Larsson J. Combined lentiviral- and RNA-mediated CRISPR/Cas9 delivery for efficient and traceable gene editing in human hematopoietic stem and progenitor cells. Sci Rep. 2020;10(1):22393.
doi pubmed - Tran NT, Graf R, Wulf-Goldenberg A, Stecklum M, Strauss G, Kuhn R, Kocks C, et al. CRISPR-Cas9-mediated ELANE mutation correction in hematopoietic stem and progenitor cells to treat severe congenital neutropenia. Mol Ther. 2020;28(12):2621-2634.
doi pubmed - Rai R, Romito M, Rivers E, Turchiano G, Blattner G, Vetharoy W, Ladon D, et al. Targeted gene correction of human hematopoietic stem cells for the treatment of Wiskott - Aldrich Syndrome. Nat Commun. 2020;11(1):4034.
doi pubmed - Rocca CJ, Rainaldi JN, Sharma J, Shi Y, Haquang JH, Luebeck J, Mali P, et al. CRISPR-Cas9 gene editing of hematopoietic stem cells from patients with Friedreich's ataxia. Mol Ther Methods Clin Dev. 2020;17:1026-1036.
doi pubmed - Goodwin M, Lee E, Lakshmanan U, Shipp S, Froessl L, Barzaghi F, Passerini L, et al. CRISPR-based gene editing enables FOXP3 gene repair in IPEX patient cells. Sci Adv. 2020;6(19):eaaz0571.
doi pubmed - Weber L, Frati G, Felix T, Hardouin G, Casini A, Wollenschlaeger C, Meneghini V, et al. Editing a gamma-globin repressor binding site restores fetal hemoglobin synthesis and corrects the sickle cell disease phenotype. Sci Adv. 2020;6(7):eaay9392.
doi pubmed - Brault J, Liu T, Bello E, Liu S, Sweeney CL, Meis RJ, Koontz S, et al. CRISPR-targeted MAGT1 insertion restores XMEN patient hematopoietic stem cells and lymphocytes. Blood. 2021;138(26):2768-2780.
doi pubmed - Sweeney CL, Pavel-Dinu M, Choi U, Brault J, Liu T, Koontz S, Li L, et al. Correction of X-CGD patient HSPCs by targeted CYBB cDNA insertion using CRISPR/Cas9 with 53BP1 inhibition for enhanced homology-directed repair. Gene Ther. 2021;28(6):373-390.
doi pubmed - Uchida N, Li L, Nassehi T, Drysdale CM, Yapundich M, Gamer J, Haro-Mora JJ, et al. Preclinical evaluation for engraftment of CD34(+) cells gene-edited at the sickle cell disease locus in xenograft mouse and non-human primate models. Cell Rep Med. 2021;2(4):100247.
doi pubmed - Psatha N, Georgakopoulou A, Li C, Nandakumar V, Georgolopoulos G, Acosta R, Paschoudi K, et al. Enhanced HbF reactivation by multiplex mutagenesis of thalassemic CD34+ cells in vitro and in vivo. Blood. 2021;138(17):1540-1553.
doi pubmed - Fananas-Baquero S, Quintana-Bustamante O, Dever DP, Alberquilla O, Sanchez-Dominguez R, Camarena J, Ojeda-Perez I, et al. Clinically relevant gene editing in hematopoietic stem cells for the treatment of pyruvate kinase deficiency. Mol Ther Methods Clin Dev. 2021;22:237-248.
doi pubmed - Li C, Georgakopoulou A, Mishra A, Gil S, Hawkins RD, Yannaki E, Lieber A. In vivo HSPC gene therapy with base editors allows for efficient reactivation of fetal gamma-globin in beta-YAC mice. Blood Adv. 2021;5(4):1122-1135.
doi pubmed - Samuelson C, Radtke S, Zhu H, Llewellyn M, Fields E, Cook S, Huang MW, et al. Multiplex CRISPR/Cas9 genome editing in hematopoietic stem cells for fetal hemoglobin reinduction generates chromosomal translocations. Mol Ther Methods Clin Dev. 2021;23:507-523.
doi pubmed - El-Kharrag R, Berckmueller KE, Madhu R, Cui M, Campoy G, Mack HM, Wolf CB, et al. Efficient polymer nanoparticle-mediated delivery of gene editing reagents into human hematopoietic stem and progenitor cells. Mol Ther. 2022;30(6):2186-2198.
doi pubmed - Karuppusamy KV, Demosthenes JP, Venkatesan V, Christopher AC, Babu P, Azhagiri MK, Jacob A, et al. The CCR5 Gene Edited CD34(+)CD90(+) Hematopoietic Stem Cell Population Serves as an Optimal Graft Source for HIV Gene Therapy. Front Immunol. 2022;13:792684.
doi pubmed - Venkatesan V, Christopher AC, Rhiel M, Azhagiri MKK, Babu P, Walavalkar K, Saravanan B, et al. Editing the core region in HPFH deletions alters fetal and adult globin expression for treatment of beta-hemoglobinopathies. Mol Ther Nucleic Acids. 2023;32:671-688.
doi pubmed - Rai R, Naseem A, Vetharoy W, Steinberg Z, Thrasher AJ, Santilli G, Cavazza A. An improved medium formulation for efficient ex vivo gene editing, expansion and engraftment of hematopoietic stem and progenitor cells. Mol Ther Methods Clin Dev. 2023;29:58-69.
doi pubmed - Wellhausen N, O'Connell RP, Lesch S, Engel NW, Rennels AK, Gonzales D, Herbst F, et al. Epitope base editing CD45 in hematopoietic cells enables universal blood cancer immune therapy. Sci Transl Med. 2023;15(714):eadi1145.
doi pubmed - Lydeard JR, Lin MI, Ge HG, Halfond A, Wang S, Jones MB, Etchin J, et al. Erratum: Development of a gene edited next-generation hematopoietic cell transplant to enable acute myeloid leukemia treatment by solving off-tumor toxicity. Mol Ther Methods Clin Dev. 2023;31:101167.
doi pubmed - Hardouin G, Antoniou P, Martinucci P, Felix T, Manceau S, Joseph L, Masson C, et al. Adenine base editor-mediated correction of the common and severe IVS1-110 (G>A) beta-thalassemia mutation. Blood. 2023;141(10):1169-1179.
doi pubmed - Brault J, Liu T, Liu S, Lawson A, Choi U, Kozhushko N, Bzhilyanskaya V, et al. CRISPR-Cas9-AAV versus lentivector transduction for genome modification of X-linked severe combined immunodeficiency hematopoietic stem cells. Front Immunol. 2022;13:1067417.
doi pubmed - Frati G, Brusson M, Sartre G, Mlayah B, Felix T, Chalumeau A, Antoniou P, et al. Safety and efficacy studies of CRISPR-Cas9 treatment of sickle cell disease highlights disease-specific responses. Mol Ther. 2024;32(12):4337-4352.
doi pubmed - Bahal S, Zinicola M, Moula SE, Whittaker TE, Schejtman A, Naseem A, Blanco E, et al. Hematopoietic stem cell gene editing rescues B-cell development in X-linked agammaglobulinemia. J Allergy Clin Immunol. 2024;154(1):195-208.e198.
doi pubmed - Demirci S, Khan MBN, Hinojosa G, Le A, Leonard A, Essawi K, Gudmundsdottir B, et al. Ex vivo culture resting time impacts transplantation outcomes of genome-edited human hematopoietic stem and progenitor cells in xenograft mouse models. Cytotherapy. 2024;26(6):641-648.
doi pubmed - Nasri M, Ritter MU, Mir P, Dannenmann B, Kaufmann MM, Arreba-Tutusaus P, Xu Y, et al. CRISPR-Cas9n-mediated ELANE promoter editing for gene therapy of severe congenital neutropenia. Mol Ther. 2024;32(6):1628-1642.
doi pubmed - Pugliano CM, Berger M, Ray RM, Sapkos K, Wu B, Laird A, Ye Y, et al. DNA-PK inhibition enhances gene editing efficiency in HSPCs for CRISPR-based treatment of X-linked hyper IgM syndrome. Mol Ther Methods Clin Dev. 2024;32(3):101297.
doi pubmed - Dudek AM, Feist WN, Sasu EJ, Luna SE, Ben-Efraim K, Bak RO, Cepika AM, et al. A simultaneous knockout knockin genome editing strategy in HSPCs potently inhibits CCR5- and CXCR4-tropic HIV-1 infection. Cell Stem Cell. 2024;31(4):499-518.e496.
doi pubmed - Park SH, Lee CM, Dever DP, Davis TH, Camarena J, Srifa W, Zhang Y, et al. Highly efficient editing of the beta-globin gene in patient-derived hematopoietic stem and progenitor cells to treat sickle cell disease. Nucleic Acids Res. 2019;47(15):7955-7972.
doi pubmed - Dever DP, Bak RO, Reinisch A, Camarena J, Washington G, Nicolas CE, Pavel-Dinu M, et al. CRISPR/Cas9 beta-globin gene targeting in human haematopoietic stem cells. Nature. 2016;539(7629):384-389.
doi pubmed - Amendola M, Brusson M, Miccio A. CRISPRthripsis: the risk of CRISPR/Cas9-induced chromothripsis in gene therapy. Stem Cells Transl Med. 2022;11(10):1003-1009.
doi pubmed - Tsuchida CA, Brandes N, Bueno R, Trinidad M, Mazumder T, Yu B, Hwang B, et al. Mitigation of chromosome loss in clinical CRISPR-Cas9-engineered T cells. Cell. 2023;186(21):4567-4582.e4520.
doi pubmed - Boutin J, Cappellen D, Rosier J, Amintas S, Dabernat S, Bedel A, Moreau-Gaudry F. ON-target adverse events of CRISPR-Cas9 nuclease: more chaotic than expected. CRISPR J. 2022;5(1):19-30.
doi pubmed - Kamminga LM, van Os R, Ausema A, Noach EJ, Weersing E, Dontje B, Vellenga E, et al. Impaired hematopoietic stem cell functioning after serial transplantation and during normal aging. Stem Cells. 2005;23(1):82-92.
doi pubmed - Akpek G, Pasquini MC, Logan B, Agovi MA, Lazarus HM, Marks DI, Bornhaeuser M, et al. Effects of spleen status on early outcomes after hematopoietic cell transplantation. Bone Marrow Transplant. 2013;48(6):825-831.
doi pubmed - Nicolini FE, Cashman JD, Hogge DE, Humphries RK, Eaves CJ. NOD/SCID mice engineered to express human IL-3, GM-CSF and Steel factor constitutively mobilize engrafted human progenitors and compromise human stem cell regeneration. Leukemia. 2004;18(2):341-347.
doi pubmed
This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, including commercial use, provided the original work is properly cited.
Journal of Hematology is published by Elmer Press Inc.